대장균 이외의 다른 생물체 용도의 클로닝 벡터

7. Cloning Vectors for Organisms Other than E. coli
대부분의 cloning 실험은 숙주로서 E. coli 와 함께 수행되며, 매우 다양한 cloning vector 들이 이 생물체에 대해 유용하다. Cloning 실험의 목적이 유전자 구조와 기능 같은 분자생물학적인 기본적 특징을 연구할 때 특히 그렇다. 그러나 어떤 경우에는 gene cloning 실험을 위해 다른 숙주를 이용하는 것이 바람직할 수도 있다. 이것은 그 목적이 유전자 연구가 아니라 중요한 대사산물(ex, hormone)의 합성을 조절하거나 향상시키고, 또는 생물체의 특징을 변화시키는(ex, 제초제 저항성 농작물에 도입시키는 등) cloning 기법을 사용하는 생물공학분야에서 특히 그렇다. 따라서 E. coli 이외의 다른 생물체를 위한 cloning vector를 생각해야 한다.
7.1 Vectors for Yeast and Other Fungi
Yeast Saccharomyces cerevisiae 는 생물공학에서 가장 중요한 생물체 중 하나이다. Yeast는 양조와 제빵에서 중요한 역할을 할 뿐만 아니라, 조작된 유전자로부터 중요한 의약품을 생산하기 위한 숙주생물체로서 이용되고 있다. Yeast를 위한 cloning vector 들의 개발은 S. cerevisiae의 대부분 균주에 존재하는 plasmid인 2㎛ circle의 발견에 의해 자극되었다.
7.1.1 Selectable markers for the 2㎛ plasmid
2㎛ circle은 크기가 6kb이고 yeast cell내에서 copy number는 70~200 사이이다. 복제는 plasmid origin을 이용하고, 복제를 위해 필요한 효소는 plasmid의 REP1, REP2 에 암호화되어 있는 단백질, 그리고 일부는 숙주세포로부터 공급받는다. 따라서 2㎛ circle은 cloning vector로서 이상적이다.

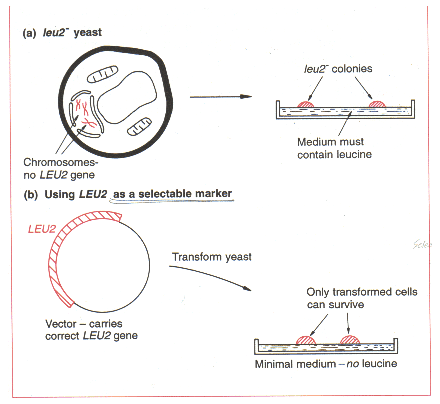
그러나 2㎛ circle 이 cloning vector로서 완벽한 것은 아니다. 우선 selectable marker가 없다. 최근의 yeast cloning vector들은 methotrexate와 copper 같은 저해제에 대해 저항성을 부여하는 유전자를 가지고 있는 것이 있긴 하지만, 대부분은 아미노산 생합성과 관련된 효소를 암호화하고 있는 yeast gene이 이용된다. 일례로 pyruvate를 leucine으로 변환시키는 효소들 중 하나인 β-isopropyl malate dehydrogenase를 암호화하고 있는 LEU2 유전자를 이용한다.
Selectable marker로써 LEU2 를 이용하기 위해선, 숙주생물이 non-functional LEU2 유전자를 가진 돌연변이체 이어야만 한다. 그러한 leu2- yeast는 leucine을 합성할 수 없고 따라서 성장배지에 leucine이 공급되는 경우에만 생존할 수 있다. Cloning 실험에서, 세포는 최소배지(추가된 아미노산이 없는)에 plating 된다. 형질전환된 세포만이 생존할 수 있고 colony를 형성할 수 있다.
2㎛ circle ; 장점 : size가 6 kb로 이상적이고, 복제수가 많다(high copynumber)
단점 : selectable marker가 존재하지 않는다.
(cf, 진핵생물에 효력이 없는 항생제 저항성 유전자를 사용할 수 없다)
7.1.2 Vectors based on the 2㎛ circle -yeast episomal plasmids
2㎛ circle로부터 유도된 vector를 yeast episomal plasmid 또는 YEp 이라 부른다. 일부 YEp은 완전한 2㎛ circle을 가지고 있고, 나머지는 단지 2㎛ 의 replication origin 만을 포함하고 있다.
- shuttle vector
- 2㎛ 의 replication origin
- selectable LEU2 gene
- entire pBR322 sequence
⇒따라서 yeast 와 E. coli 모두에서 복제할 수 있고 선별될 수 있다.
host cell로서 leu2- mutant를 이용
leu2 2㎛ circle을 leu2- mutant에 형질전환 시킴
-grow at no leucine medium → transformant
-can not grow no leucine medium → non-transformant

Shuttle vector를 사용하는 이유는 형질전환된 yeast colony로부터 재조합 DNA분자를 회수하기 어려울 수도 있기 때문이다. 이러한 문제는 yeast vector가 yeast chromosome 내부로 통합되는 경우 발생하며, 이 경우 재조합 DNA분자의 순수분리는 불가능하다. 따라서 yeast에서 cloning 실험을 할 때, 표준적인 실험방법은 처음에 cloning 실험을 E. coli 와 함께 수행하여 recombinant를 선별하고 난 후, recombinant plasmid를 분리하여 특징을 결정하고, yeast 내부로 그 correct molecule을 도입시키는 것이다.

7.1.3 A YEp may insert into yeast chromosomal DNA
'Episomal' 이라는 말은 YEp 이 독립적인 plasmid 로서 복제할 수 있다는 것을 가르킬 뿐만 아니라 yeast chromosome 의 한 곳으로 통합될 수 있다는 것을 암시하는 것이다. 통합은 selectable marker 로서 vector에 존재하는 유전자가 yeast chromosome에 존재하는 돌연변이 유전자와 매우 가까운 상동성인 경우 일어난다. 예를 들어 YEp13에서, 재조합은 plasmid LEU2 유전자와 yeast mutant LEU2 사이에서 일어날 수 있다. 그 결과 전체 plasmid가 yeast chromosome의 한 부위 내부로 삽입된다. 그 plasmid는 통합된 상태로 유지될 수도 있고 또는 나중에 재조합이 일어나서 다시 excision되기도 한다.

7.1.4 Other types of yeast cloning vector
1) Yeast integrative plasmids(YIps)
기본적으로 yeast gene을 가진 bacterial plasmid URA3 gene이 삽입된 pBR322.
이 유전자는 pyrimidine nucleotide 생합성 단계 중 하나를 촉매하는 효소인 orotidine-5'-phosphate decarboxylase를 암호화하고 있으며 LEU2 와 동일한 방식으로 selectable marker로써 이용됨.
YIp 은 2㎛ circle의 어떠한 부분도 가지고 있지 않기 때문에 plasmid 처럼 복제할 수 없으며 그것의 복제는 yeast chromosome 내부로의 통합에 의존함.
2) Yeast replicative plasmids(YRps)
Replication origin을 포함하는 chromosomal DNA 서열을 가지고 있기 때문에 독립적인 plasmid로서 복제할 수 있음.
Replication origin은 selectable marker로 이용될 수 있는 몇 개의 yeast gene과 매우 가까운 위치에 있는 것으로 알려져 있음.
Yeast gene TRP1 이 삽입된 pBR322.
Tryptophan 생합성과 관련된 이 유전자는 chromosome의 replication origin과 인접해 있다. YRp7 에 존재하는 yeast DNA 단편은 TRP1 과 그 origin을 모두 가지고 있다.

특정 cloning 실험을 위해 어떤 유형의 yeast vector가 가장 적합한가를 결정할 때 세가지 측면에서 고려해야 한다.
1) Transformation frequency : cloned gene을 얻고자 하는 경우
YEp : 10,000~100,000 transformed cell /㎍
YRp : 1,000~10,000 transformed cell /㎍
YIp : 1,000 transformants /㎍ 이하
2) Copy number : cloned gene 으로부터 단백질을 얻고자 하는 경우
YEp : 20~50 / cell
YRp : 5~100 / cell
YIp : 1 / cell
3) Stability : 배양액 내에 오랜기간 동안 cloned gene을 보존해야 하는 경우
YEp : unstable
YRp : extremely unstable
YIp : most stable
7.1.5 Artificial chromosomes can be used to clone huge pieces of DNA in yeast
Yeast cloning vector 의 마지막 유형은, gene cloning 에 대한 완전히 새로운 접근 방식으로써, yeast artificial chromosome을 의미하는 YAC 이다. YAC 의 개발은 진핵생물의 염색체 구조에 대한 기초적인 연구로부터 파생된 것이었다.
염색체 구조가 밝혀짐으로해서 개별적인 구성요소(centromere, telomere, origins)들이 재조합 DNA 기술에 의해 분리되고, 그 후 시험관 내에서 다시 서로 결합되어 인공염색체가 창조될 가능성이 높아졌다. 효모염색체에 존재하는 DNA분자가 수백 kb이기 때문에 인공염색체를 이용하면 어떤 다른 유형의 vector가 가질 수 있는 것보다 훨씬 더 큰 DNA단편을 cloning하는 것이 가능할 수도 있다.
(a) The structure and use of a YAC vector

AC vector의 전형적인 예는 pYAC3이다. pYAC3는 얼핏보면 인공염색체처럼 보이지 않지만, 자세히 관찰해보면 그것의 특징들을 가지고 있다.
pYAC3는 수많은 yeast gene이 삽입되어 있는 pBR322이다. 이들 유전자 중 두 유전자 URA3 와 TRP1 은 각각 YIp5와 YRp7에서 selectable marker 로써 이미 소개되었다. YRp7에서와 같이, TRP1 을 가지고 있는 DNA단편은 또한 replication origin을 가지고 있지만, pYAC3에서 이 단편은 CEN4 라는 서열을 포함하고 있어서 더 연장되어 있다. CEN4 는 chromosome 4의 centromere 부위로부터 얻어진 DNA이다. 그러므로 TRP1-origin-CEN4 단편은 인공염색체의 세가지 구성요소중 두가지를 가지고 있는 것이다.
세 번째 구성요소인 telomere는 TEL 이라 불리우는 두 개의 서열에 의해 제공된다. 이들은 그들 자체로 완전한 telomere서열은 아니지만, 일단 yeast 핵내에서 그들은 telomere가 형성될 seeding sequence 역할을 한다. 나머지 한 부분은 SUP4 이며 이것은 cloning 실험동안 새로운 DNA가 삽입되는 selectable marker이다.
pYAC3를 이용한 cloning 방법은 다음과 같다.
- 우선 BamHI과 SnaBI을 사용하여 double digestion시킨다.
- BamHI fragment를 제거하면 양 말단에 TEL sequence와 SnaBI site가 존재하는 두 개의 arm이 남는다.
- SnaBI(TACGTA를 인식하여 blun end를 만드는 제한효소)절단 부위처럼 blunt end를 가진 조작될 DNA를 두 arm 사이에 ligation 시켜 인공염색체를 만든다.
- Protoplast transformation 방법을 이용하여 인공염색체를 S. cerevisiae 내로 도입시킨다. 이때 사용되는 효모균주는 trp1-ura3- 인 double mutant로서 인공염색체 상의 두가지 marker에 의해 trp1+ura3+ 로 전환된다.
- 따라서 형질전환체는 정확히 제조된 인공염색체를 가진 세포만 성장할 수 있는 최소배지에 plating함으로써 선별된다.
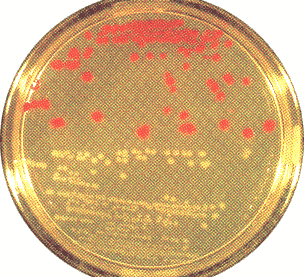
- Vector 내에 삽입 DNA의 존재는 SUP4의 insertional inactivation을 검사항으로써 확인될 수 있다. 이것은 간단한 color test에 의해 수행된다. White colony는 recombinant이고 red colony는 아니다.
(b) Applications for YAC vectors
일부 포유동물의 유전자들은 길이가 100kb 보다 훨씬 크기 때문에, 대부분의 E. coli cloning system의 능력 이상이지만 YAC vector의 범위에는 포함된다. 따라서 YAC은 재조합 DNA 기술에 의한 분석을 위해 이전에 다룰 수 없었던 유전자들의 기능과 발현양상을 연구하기 위한 길을 열어준 것이다. 이러한 새로운 차원의 실험기술은 유전자가 정상적으로 존재하고 잇는 생물체내에서 수행되는 기능적인 분석을 가능하게 하는 포유동물 세포에서 YAC이 증식될 수 있다는 발견에 의해 최근에 수행되고 있다.
YAC은 gene library의 생산에서도 똑같이 중요하다. YAC vector는 보통 600kb 단편을 cloning하기 위해 이용되며, 특별한 유형은 1400kb 이상의 DNA를 조작할 수 있다. 후자의 경우를 이용하면 인간의 gene library를 단지 6500 clone 으로 줄일 수 있다.
7.1.6 Vectors for other yeasts and fungi
Yeast 와 fungi 의 다른 종들을 위한 cloning vector는 이들 생물체들의 기본적인 분자 생물학적 연구를 위해 그리고 생물공학에서 yeast 와 fungi 의 이용가능성을 확장시키기 위해 필요하다. 2㎛ circle에 기초한 episomal plasmid는 극소수의 다른 yeast 유형에서 복제할 수 있지만 종의 범위는 폭넓지 않다.
어떤 경우에, 생물공학에서 YIp과 같은 integrative plasmid가 더욱 적합하다. 왜냐하면 이들은 생물반응체 내에서 장기간동안 성장할 수 있는 안정적인 재조합체를 제공하기 때문이다. 효율적인 integrative vector들은 현재 Pichia pastoris, Kluveromyces lactis 같은 yeast와 Aspergillus nidulans, Neurospora crassa 같은 filamentous fungi를 포함하는 수많은 종들을 위해 유용하다.
7.2 Cloning Vectors for Higher Plants
숙주 생물체로써 고등식물을 이용하는 gene cloning에는 중요한 잠재적 이점이 있다. 이미 gene cloning을 통해 virus와 곤충에 저항성을 가진 실험 식물이 만들어지고 있으며, 저항성을 향상시킨 유전학적으로 조작된 tomato는 supermarket에 공급되고 있다.
고등식물을 위한 세가지 유형의 vector system이 사용되고 있다.
① Tumor-inducing(Ti) plasmid of Agrobacterium tumefaciens
Agrobacterium에 자연적으로 존재하는 plasmid에 기초한 vector
② Direct gene transfer to protoplast of plant cell
Plant cloning vector에 부착되지 않은 DNA단편을 사용하는 direct gene transfer
③ Plant virus vector
Plant virus에 기초한 vector
7.2.1 Agrobacterium tumefaciens-nature's smallest genetic engineer
Agrobacterium tumefaciens는 많은 쌍자엽 식물에 crown gall disease를 일으키는 토양미생물이다. Crown gall은 상처난 줄기에 Agrobacterium tumefaciens가 침입할 때 발생한다. 감염 후에 이 세균은 줄기조직의 암 같은 증식을 유발시킨다.

Crown gall disease를 유발시키는 능력은 이 세균내에 Ti(Tumor Inducing) plasmid의 존재와 관련이 있다. 이것은 200 kb 이상의 거대한 plasmid이며 감염과정과 관련된 많은 유전자들을 가지고 있다.
Ti plasmid의 한 가지 주목할 만한 특징은 감염 후에 그 일부가 plant chromosomal DNA 내부로 통합된다는 것이다. 이 부위를 T(transfer)-DNA라 부르며, 균주에 따라 그 크기는 15~30 kb 사이이다. 그것은 식물세포내에서 안정적인 형태로 유지되어, 염색체의 통합부위로써 딸세포로 전달된다. 그러나 가장 놀랄 만한 특징은 그 T-DNA가 식물세포내에서 발현되는 8개의 유전자를 가지고 있으며 형질전환된 세포의 암적인 특징을 담당하고 있다는 것이다. 또한 이들 유전자는 그 세균이 영양분으로써 이용하는 opine 이라 불리우는 비정상적인 화합물을 직접 합성한다. 짧게 말해서, Agrobacterium tumefaciens는 자신의 목적을 위해 식물세포를 유전학적으로 조작한다.

① Plant cell의 물리적인 상처
② phenolic compound 분비
③ A. tumefaciens가 phenolic compound인식
④ T-DNA 이동
⑤ 숙주 chromosome에 통합
⑥ Opine 합성
* T-DNA 대신 유용한 gene을 삽입(ex, human hormone gene, vitamin gene, vaccine gene)
* 단점 : 숙주가 쌍자엽 식물로 한정되어 있을 뿐 아니라 숙주 특이성이 있다.
(a) Using the Ti plasmid to introduce new genes into a plant cell
Ti plasmid를 이용하기 위해 필요한 것은 T-DNA내에 새로운 유전자를 삽입시키는 것이다. 그러면 그 세균이 plant chromosomal DNA내로 그들을 통합시키는 어려운 일을 해줄 것이다. 그러나 실제로는 Ti plasmid의 거대한 크기가 조작을 매우 어렵게 만들기 때문에 매우 까다로운 일이다. 물론 주요한 문제점은 유일한 제한효소부위가 없다는 것이다. 새로운 DNA를 plasmid내로 삽입시키기 위해선 새로운 방법이 개발되어야만 한다.
1) The binary vector strategy
이 방법은 T-DNA가 Ti plasmid의 나머지 부분과 물리적으로 부착되어 있어야 할 필요가 없다는 사실에 기초한 것이다. 비교적 작은 분자상에 T-DNA를 가지고 있는 것과 그 plasmid의 나머지 부분을 가진 형태의 two-plasmid system은 식물세포를 형질전환시키는데 효과적이다. 실제로 Agrobacterium tumefaciens의 일부 균주와 관련 Agrobacteria는 자연적인 binary plasmid system을 가지고 있다. T-DNA plasmid는 유일한 제한 효소부위를 가질 만큼, 또 표준적인 기술을 사용하여 조작될 만큼 충분히 작다.
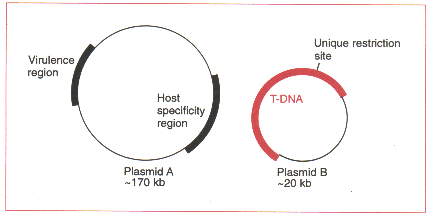
2) The cointegration strategy
이 방법은 pBR322 또는 유사한 E. coli vector에 기초하였지만, T-DNA의 작은 부분만을 가지고 있는 완전히 새로운 plasmid를 사용한다.
새로운 분자와 Ti plasmid 사이의 homology는 두 분자가 동일한 Agrobacterium tumefaciens내네 존재할 경우, 재조합에 의해 T-DNA부위 내부로 pBR plasmid가 통합될 수 있다는 것을 의미한다. 따라서 cloning될 유전자를 작은 pBR plasmid상에 특정 제한효소부위내로 삽입시키고, Ti plasmid를 가진 Agrobacterium tumefaciens안으로 도입시키면, 자연적인 재조합과정에 의해 T-DNA 내부로 새로운 유전자가 통합된다. 이 후 식물에 감염시키면 T-DNA와 함께 새로운 유전자가 식물 염색체 내부에 삽입된다.

(b) Production of transformed plants with the Ti plasmid
조작되어진 Ti plasmid를 가진 Agrobacterium tumefaciens가 자연적인 방법으로 식물내로 도입된다면 crown gall을 유발시킬 세포들만 조작된 유전자를 가지게 될 것이다. 이것은 분명히 생물공학자에게 가치가 없다. 대신 식물의 모든 세포내로 새로운 유전자를 도입시키는 방법이 필요하다. 가장 간단한 방법은 성숙한 식물이 아닌 배양중인 식물세포나 protoplast에 감염시키는 것이다.

세포벽이 재형성되고 있는 식물세포와 protoplast는 미생물처럼 동일한 방법으로 처리될 수 있다. 형질전환된 세포로부터 재형성된 성숙한 식물은 모든 세포에 조작된 유전자를 가지고 있을 것이며 조작된 유전자를 그 자손세대로 전달할 것이다.
그러나 형질전환된 식물의 재형성은 Ti vector의 병원성이 해제되어 그 형질전환된 세포가 암의 특징을 나타내지 않는 경우에만 일어날 것이다. Disarming은, cancer 유전자들이 모두 T-DNA에 위치하고 있고 감염과정을 위해 필요치 않으며, 감염성은 Ti plasmid의 virulence region에 의해 통제되기 때문에 가능하다. 사실 감염에 관련된 T-DNA의 부분은 단지 식물 DNA에 통합된 부위의 왼쪽과 오른쪽 경계에서 발견되는 두 개의 25bp 반복서열 뿐이다. 따라서 정상적인 T-DNA로부터 모든 cancer유전자를 제거시키는 것이 가능하며 감염과정의 손상없이 그들을 완전히 새로운 유전자들로 대치시킬 수 있다.
수많은 disarmed Ti cloning vector가 현재 유용하며 전형적인 예로 binary vector pBIN19를 들 수 있다. 이 vector는 T-DNA의 왼쪽과 오른쪽 경계부위, lacZ' gene, kanamycin 저항성 유전자를 가지고 있다. Yeast shuttle vector와 마찬가지로, pBIN19내로 조작될 유전자를 삽입하는 처음 조작은 E. coli 에서 수행한다. 그리고 나서 recombinant pBIN19를 Agrobacterium tumefaciens로, 그 후에 식물로 이동시키다. 형질전환된 식물세포는 kanamycin을 함유한 agar medium에 plating함으로써 선별된다.

pBIN19같은 Ti vector들은 최근에 Agrobacterium rhizogenes의 Ri plasmid에 기초한 관련 vector들에 의해 보완되고 있다. Ri plasmid와 Ti plasmid는 매우 유사하며, 주된 차이점은 Ri plasmid로부터 식물로의 T-DNA의 이동은 crown gall이 아닌 hairy root diseast를 유발시킨다는 점이다.
7.2.2 Cloning gene in plants by direct gene transfer
Direct gene transfer는 supercoiled bacterial plasmid가 식물 염색체중 한 개의 내부로 재조합에 의해 통합될 수도 있다는 관찰에 기초하고 있다. 이 재조합 기작에 대해선 잘 알려져 있지 않다. 사실 통합은 식물 염색체중 어느 한 염색체 내부에 어떤 위치에 무작위적으로 일어난다.

Selectable marker와 조작될 유전자를 가진 supercoiled plasmid DNA를 어떻게 식물세포 내부로 도입시킬 수 있는가?
1) Original method
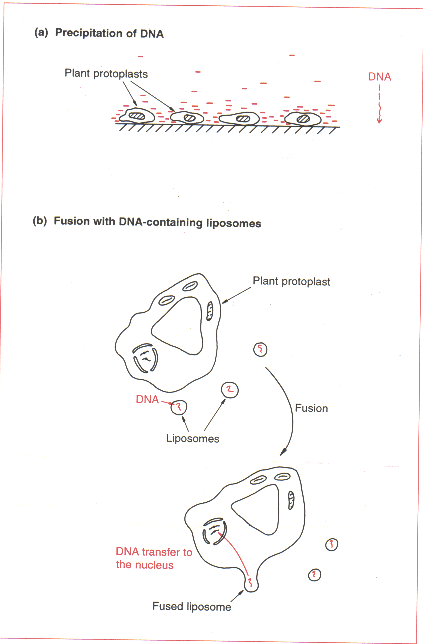
Polyethylene glycol, polymeric negatively-charged compound로 구성된 점착성 용액에 재현탁된 protoplast를 이용하는 방법으로, protoplast표면에 DNA를 침전시키고 endocytosis에 의해 uptake를 유도시킨다고 생각되고 있다.
2) More efficient method
Electroporation, microinjection, 그리고 DNA-containing liposome을 이용한 융합법 등이 최근 이용되고 있다. 처리 후에는 세포벽을 재생성시키기 위해 protoplast를 며칠 간 용액속에 방치시킨다. 그 다음에 형질전환체를 동정하기 위해 선택배지에 plating하고 callus를 완전한 식물이 성장될 수 있는 배지로 옮긴다.
* 단점 : regeneration이 어렵다.
* 장점 : 숙주특이성이 없다.
7.2.3 Attempts to use plant viruses as cloning vectors
대부분의 식물들은 virus에 의해 감염된다. 그렇다면 식물에서 유전자를 조작하기 위해 virus가 이용될 수 있을까? 한가지 문제점은 주요한 식물 virus가 RNA genome을 가지고 있다는 점이다. RNA는 조작하기가 어렵기 때문에 RNA virus는 cloning vector로써 유용하지 못하다. DNA virus중 단지 두 종류가 고등식물에 감염하는 것으로 알려져 있다(caulimoviruses and geminiviruses).
1) Caulimovirus vector는 두 가지 단점을 가지고 있다.
① Size limitation
Non-essential section을 제거시킨 후에도 삽입될 수 있는 DNA크기는 여전히 매우 제한적이다(0.2~0.3kb)
② Narrow host range
숙주 범위가 매우 좁아서 단지 순무, 양배추, 꽃양배추에만 조작실험이 가능하다.
2) Geminivirus
옥수수와 밀 같은 중요한 작물에 자연적으로 감염하기 때문에 얼핏보면 유용할 것 같지만, infection cycle동안에 genome의 재배열과 결손이 일어나기 때문에 삽입된 DNA에 어떤 추가적인 DNA가 뒤섞일 수 있다. 또한 이 virus는 작물들에게 감염으로 인한 피해를 일으킬 수 있기 때문에 geminivirus cloning vector의 사용은, 숙주로부터 빠져나온 vector의 위험성과 자연상의 식물군집에 감염되는 것을 피하기 위해 엄격한 통제하에서 수행되어야 한다.
7.2.4 Problems with cloning genes in monocotyledonous plants
첫 번째 어려움은 자연상에서 A. tumefaciens 와 A. rhizogenes 가 쌍자엽 식물에만 감염한다는 사실에 있다.
Direct gene transfer 역시 아직은 불완전한 해결책이다.
7.3 Cloning Vectors for Animal Cells
동물세포내에서 유전자를 조작하기 위한 vector system을 개발하기 위해 상당한 노력을 하고 있다. 이 vector는 E. coli 또는 yeast에서 조작될 경우, 정상적으로 기능하지 못하는 유전자로부터 단백질을 합성하기 위해 생물공학에서 필요로 하고 있으며, 조작기술에 의해 환자에게 정상적인 유전자를 도입시켜 돌연변이 유전자를 대체시킴으로써 유전질환을 고치는 유전자 치료법을 위한 치료방법을 고안하기 위해서도 필요하다.
Direct gene transfer가 가능하지만 그 효과는 매우 낮다. YAC과 동일한 개념으로, 포유동물의 인공염색체도 또한 가능하긴 하지만 실제로 보고된 바는 아직 없다. 따라서 대부분의 조작실험은 virus에 기초한 vector에 의존하고 있다.
7.3.1 Vectors based on animal viruses
동물세포와 관련된 최초의 조작실험은 SV40에 기초한 vector를 이용하여 1979년에 수행되었다. 이 virus는 일부 포유동물에 감염하여, 어떤 숙주에서는 lytic cycle을, 나머지에서는 lysogenic cycle을 진행시킨다. SV40의 문제점은 size limitation이며, 따라서 조작될 DNA를 삽입시키기 위해 기존의 유전자를 새로운 DNA로 대체시켜야 한다.

1979년 이후, 다른 유형의 수많은 virus가 동물에서 유전자를 조작하기 위해 사용되어 왔다. Adenovirus는 SV40 vector보다 더 큰 단편을 삽입시킬 수 있고 papillomavirus도 마찬가지이며 이 virus는 또한 안정적인 형질전환된 세포를 얻을 수 있다는 장점이 있다.
많은 포유동물 virus는 감염후에 곧 숙주세포를 죽게 하기 때문에, 단기간 형질전환 실험이 아닌 경우에는 특별한 방법이 사용되어야 한다. 소에 혹을 발생시키는 bovine papillomavirus(BPV)는 쥐 세포에서 비정상적인 infection cycle을 가지게 되어, 세포 당 약 100분자의 plasmid를 형성한다. 또 쥐 세포를 사멸시키지 않으며 BPV분자는 숙주의 세포분열을 통해 딸세포로 전달된다. BPV와 pBR322서열로 구성된 shuttle vector는 쥐와 세균에서 모두 복제할 수 있기 때문에 동물세포를 이용하는 생물공학에서 매우 가치가 높다. 또한 곤충세포내에서 조작된 유전자로부터 많은 양의 단백질을 얻을 수 있는 baculovirus도 유용하다.